

Ismael Gárate-Lizárraga, Gerardo Verdugo-Díaz y Rogelio González-Armas
Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas (CICIMAR), Av. Instituto Politécnico Nacional s/n, Col. Playa Palo de Santa Rita, La Paz, Baja California Sur, C.23096, México
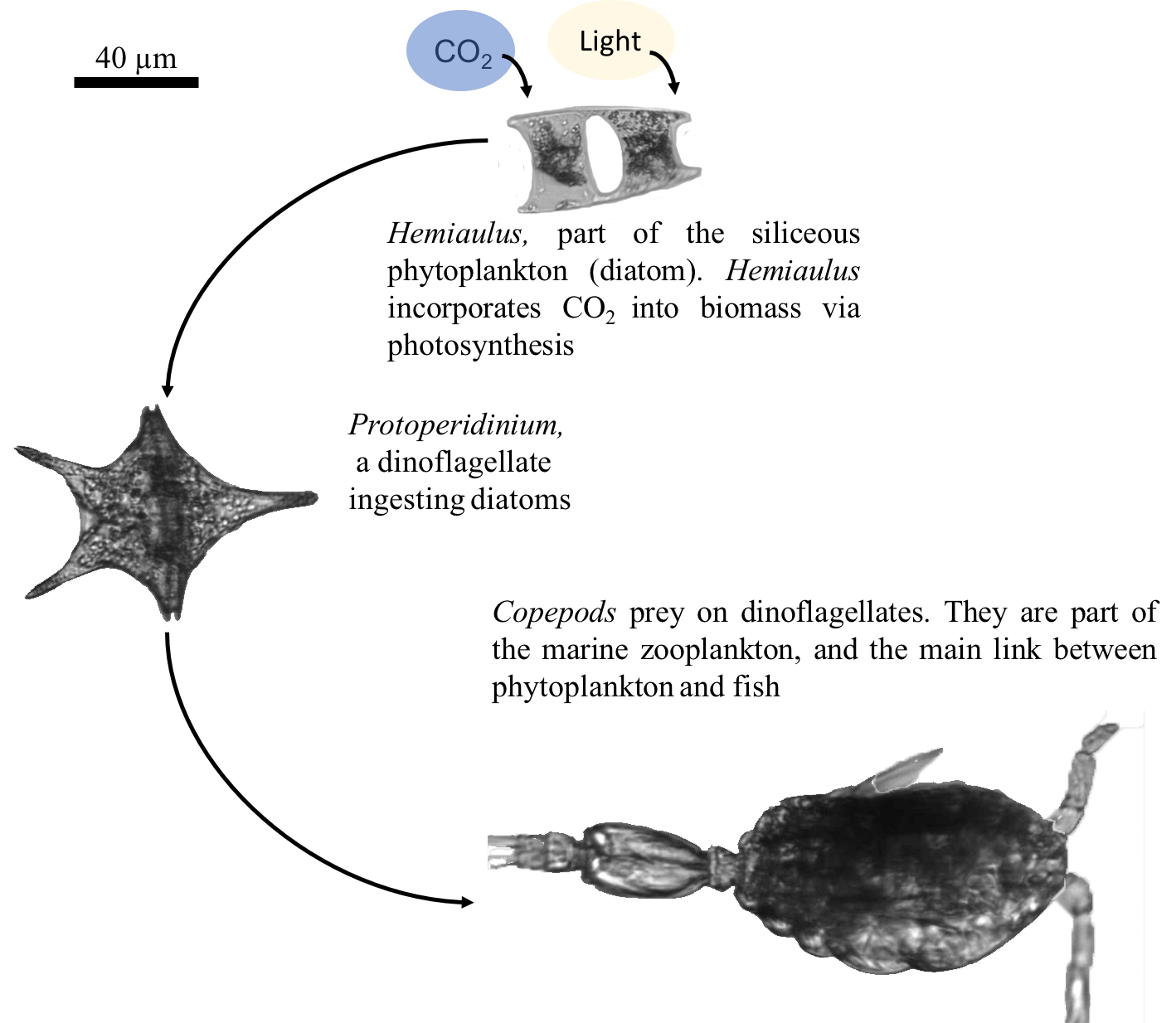
El término «plancton» fue acuñado por primera vez por el biólogo alemán Victor Hensen en 1887. La palabra «plancton» proviene del griego «planktos», que significa «a la deriva», o errante y se utiliza para describir a los organismos que van a la deriva en la superficie del agua o bien, cerca de ella y que no son capaces de desplazarse en contra de las corrientes marinas. No obstante, muchos organismos planctónicos, pueden moverse a lo largo de la columna de agua. El plancton se distribuye en casi todos los ecosistemas acuáticos y juega un papel importante en las redes tróficas basadas en el modelo clásico nutriente-fitoplancton-zooplancton (Fig. 1).

Figura 1. Modelo clásico simplificado de una red trófica (https://schmidtocean.org/wp-content/uploads/Food-web.jpg).
{kind=link}
El plancton, además de su naturaleza y de su tamaño, puede subdividirse en consideración de los grupos funcionales que lo conforman: autótrofos, heterótrofos y mixótrofos. El plancton autótrofo o fitoplancton está compuesto por organismos microscópicos unicelulares, o bien que forman cadenas y que realizan fotosíntesis, por lo cual están restringidos a vivir desde la superficie hasta la profundidad a la cual puede llegar la luz solar en suficiente cantidad para activar el proceso fotosintético. Se consideran autótrofos porque producen su propia reserva energética, necesaria para sobrevivir, producir nuevas estructuras y crecer. Son conocidos como los productores primarios en la cadena trófica, ya que son capaces de convertir el dióxido de carbono, y la energía del sol en materia orgánica utilizable por organismos de otros niveles tróficos superiores.
El fitoplancton es crucial en el ciclo del carbono, ya que puede aportar el 50% de toda la actividad fotosintética y de la producción de oxígeno (Jahan y Singh, 2023). Si bien, la fotoautotrofía es el principal modo trófico entre estos organismos, la mixotrofía es una estrategia de alimentación que presentan una fracción considerable de los componentes del fitoplancton, los cuales pueden nutrirse a través de la fotosíntesis y, al mismo tiempo, capturando a sus presas. Esta capacidad se considera una ventaja competitiva de algunas microalgas en situaciones de estrés, generadas por déficit de luz o de nutrientes en el medio (Jones 2000, Medina-Sánchez et al., 2004). Mitra et al. (2016) definieron la mixotrofía como la capacidad de realizar foto y fagotrofía por una sola célula, ya sea de manera obligada o facultativa. Stoecker et al. (2017) refieren que la mixotrofía puede considerar otras formas alternativas de nutrición y de adquisición de carbono (C), así como también, que puede incluir la adquisición de nitrógeno (N), fósforo (P), oligoelementos, hierro (Fe), nutrientes traza (vitaminas) y energía.
Los organismos mixótrofos más conocidos y de mayor distribución son flagelados unicelulares pequeños, con cloroplastos funcionales que ingieren partículas mediante fagotrofía (Stoecker, 1998). Se ha sugerido que la mixotrofía contribuye a un aumento del ~35% del flujo de carbono hacia organismos de mayor tamaño, ejerciendo una influencia sustancial en el ciclo global del carbono (Kumar et al., 2024). Dentro de los grupos funcionales del fitoplancton, las diatomeas son conocidas por su gran variabilidad y elevada productividad primaria, siendo responsables de alrededor del 20% de la fijación anual de carbono a nivel mundial (Villanova y Peteal, 2023) y presentan flexibilidad trófica que les permite, en determinadas condiciones crecer de forma mixotrófica. Hasta ahora, sólo se ha comprobado el metabolismo mixotrófico de pocas especies de diatomeas (ver Kumar et al., 2024). La figura 2 muestra la presencia de diatomeas autótrofas, con un cloroplasto (Fig. 2a), dos (Fig. 2b) o cuantiosos cloroplastos (Figs. 2c-i).
Los dinoflagelados presentan una gran diversidad de estilos de vida y modos tróficos, y se han dividido en especies autótrofas y heterótrofas. Sin embargo, ahora se sabe que esa clasificación no es adecuada, ya que muchas especies pueden combinar la fototrofía y la captación de presas, y ser así mixotróficas (Hansen y Tilmann, 2020). Sin embargo, la presencia de «partículas» en el interior de un dinoflagelado o de otras microalgas no demuestra necesariamente la existencia de fagotrofia, sino que pueden ser productos de almacenamiento interno o de bacterias endosimbióticas (Figs. 2j-C). Algunas inclusiones pueden implicar infecciones endoparasitarias (Jacobson, 1999).

Figura 2. Microalgas de la Bahía de La Paz que presentan diferentes estrategias de alimentación: a) Chaetoceros tortissimus, b) Meuniera membranacea, c) Trieres mobiliensis d) Stephanopysis palmeriama, e) Asteromphalus heptactis, f) A. flabellatus g) A. arachne, h) Licmophora abbreviata, i) Actinocyclus cuneiformis, j) Akashiwo sanguinea, k) Gymnodinium catenatum, l) Gymnodinium amphora, m) Cucumeridinium coeruleum, n) Balechina pachydermata, ñ) Spatulodinium pseudonoctiluca, o) Dinophysis fortii, p) Phalacroma doryphorum, q) Phalacroma oxytoxoides, r) Podolampas bipes, s) Blepharocysta sp., t) Protoperidinium sp., u) Tripos furca, v) Prorocentrum concavum, w) Gambierdiscus sp., x) Sinophysis canaliculata, y) Octactis pulchra, z) Dictyocha californica, A) Hermesinum adriaticum, B) Ebria tripartita, C) 2 vistas de Eutreptiella sp.
Los dinoflagelados fotosintéticos se han estudiado intensamente debido a que suelen formar florecimientos en regiones costeras y por su papel en la productividad primaria marina (Buskey et al., 1994). En hábitats oligotróficos la mixotrofia se considera un importante modo de nutrición (Jones, 1994, 2000). Como ya mencionamos, los dinoflagelados heterótrofos desempeñan un papel importante en el ciclo del carbono y el flujo de energía en la comunidad planctónica marina. En la Bahía de La Paz, se han observado algunas especies mixotróficas de dinoflagelados desnudos como Akashiwo sanguinea y Gymnodinium catenatum (Figs. 2j-2k) y heterótrofos como Gymnodinium amphora, Cucumeridinium coeruleum, Balechina pachydermata, y Spatulodinium pseudonoctiluca (Figs. 2l, 2m, 2n y 2ñ), suelen ser frecuentes durante los eventos de mareas rojas.
Un proceso simbiótico común en algunos dinoflagelados es la cleptoplastía, en el cual, los cloroplastos de otras microalgas son secuestrados por el hospedero. Los dinoflagelados que secuestran exclusivamente cloroplastos de sus presas pertenecen a los géneros Dinophysis y Phalacroma (Figs. 2o-2q). Las especies de Dinophysis son mixótrofas, ya que combinan la fototrofia, por medio de cleptoplastidios robados a sus presas. con una fagotrofía muy selectiva sobre el ciliado Mesodinium. No obstante, otras especies de dinoflagelados que utilizan la cleptoplastía son Amylax triacantha y A. triacantha var. buxus, los cuales ingieren criptofíceas. Recientemente Gárate-Lizárraga y Okoldokov (2022) reportaron que los taxa Podolampas bipes (Fig. 2r), P. elegans. P. palmipes, P. reticulata y P. spinifera presentaron kleptoplastos endocitobiontes, tentativamente de la clase Dictyochophyceae. También reportaron a Blepharocysta splendor–maris (Fig. 2s) en su primer estadio de alimentación. La mixotrofia también se ha observado en dinoflagelados planctónicos (Figs. 2o-2u) como bentónicos Prorocentrum, Gambierdiscus y Sinophysis (Figs. 2v, 2w y 2x).
Los silicoflagelados y los ebridios, al igual que las diatomeas, forman parte del fitoplancton marino que segregan sílice. Los primeros son fotosintéticos y heterótrofos unicelulares presentes en ambientes marinos que se caracterizan por presentar un esqueleto silíceo integrado por una red de barras y espinas (Figs. 2y y 2z). Los ebridios, también presentan un endoesqueleto y son más comunes en aguas templadas y frías y no poseen cromatóforos, por lo que son heterótrofos, siendo Hermesinium adriaticum (Fig. 2A) y Ebria tripartita (Fig. 2B) las especies más frecuentes en la Bahía de La Paz. Las euglenofitas han sido poco estudiado en ambientes marinos y también se caracterizan por tener diversos modos de nutrición, como fototrofía y mixotrofía (Fig. 2C).
No obstante, de su relevancia, la mixotrofía y la fagotrofía, continúan siendo poco conocidas o bien, ignoradas y son pocos los estudios que muestran su impacto sobre el resto de los integrantes de las redes tróficas marinas y sobre la producción de recursos en el océano. En México se han realizado pocos estudios donde se aborda la forma de nutrición de las especies del fitoplancton, enfocándose principalmente al estudio de las comunidades o de las especies formadoras de mareas rojas. Es en este tipo de eventos, donde se han observado estas estrategias nutricionales, principalmente en los del dinoflagelado Noctiluca scintillans y el ciliado Mesodinium rubrum (Gárate-Lizárraga et al., 2001). N. scintillans es un voraz consumidor de diatomeas y de huevos de copépodos, mientras que M. rubrum ingiere microalgas del grupo de las criptofitas, funcionado como un verdadero fotótrofo, con la capacidad de regular y dividir los cloroplastos (Johnson et al., 2017). Gárate-Lizárraga et al. (2002) y López-Cortés et al. (2008) realizaron el perfil pigmentario de M. rubrum en muestras del Golfo de California y encontraron la presencia aloxantina, pigmento característico de las criptofíceas y alimento principal de este ciliado, y que además retiene sus plastidios y mitocondrias. De acuerdo con lo anterior, consideramos necesario que se desarrollen estudios que aborden diferentes aspectos de las estrategias de alimentación de los grupos del fitoplancton, con el fin de conocer que especies utilizan una determinada estrategia, en qué condiciones ocurren, y que implicaciones ecológicas pueden presentar para los organismos de los niveles tróficos superiores.
Referencias
Buskey EJ, Coulter CJ, & Brown SL. (1994) Feeding, growth and bioluminescence of the heterotrophic dinoflagellate Protoperidinium huberi. Marine Biology, 121:373-380.
Gárate-Lizárraga I, & Okolodkov YB. (2022). The family Podolampadaceae (Dinoflagellata) in Mexican waters. Hidrobiológica, 32(3), 251-263.
Gárate-Lizárraga I, Hernández-Orozco ML, Band-Schmidt CJ, Serrano-Casillas G. (2001). Red Tides along the coasts of Baja California Sur, México (1984-2001). Oceánides, 16(2), 127-134.
Gárate-Lizárraga I, Band-Schmidt, CJ, Cervantes-Duarte R, & Escobedo-Urías D. (2002). Mareas rojas de Mesodinium rubrum (Lohmann) Hamburger y Buddenbrock en el Golfo de California (invierno de 1998). Hidrobiológica, 12(1), 15-20.
Hansen PJ, & Tillmann U. (2020). Mixotrophy in dinoflagellates: Prey Selection, Physiology and Ecological Importance. In S. R. V. Durvasula (Ed.), Dinoflagellates: Classification, Evolution, Physiology and Ecological Significance (pp. 201-260). Nova Science Publishers.
Jahan S. (2023). The Role of phytoplankton in the environment and in human life, a Review. Basra Journal of Science. 41, 392-411.
Jones RI. (2000). Mixotrophy in planktonic protists: an overview. Freshwater Biology 45:219-226.
Johnson MD, Oldach D, Delwiche CF, & Stoecker DK. (2007) Retention of transcriptionally active cryptophyte nuclei by the ciliate Myrionecta rubra. Nature 445(7126):426–428.
López-Cortés, DJ, Gárate-Lizárraga I, Bustillos-Guzmán JJ, & Hernández-Sandoval FE. (2008). Blooms of Myrionecta rubra in Bahía de La Paz, Gulf of California, during early summer of 2005. CICIMAR Oceánides, 1-10.
Medina-Sánchez JM, Villar-Argaiz M, & Carrillo PC. (2004). Neither with nor without you: A complex algal control on bacterioplankton in a high mountain lake. Limnology and Oceanography, 49, 1722-1733.
Mitra A, Flynn KJ, Tillmann U, Raven JA, Caron DA, Stoecker DK, Not F, Hansen PJ, et al. (2016). Defining planktonic Protist functional groups on mechanisms for energy and nutrient acquisition: incorporation of diverse Mixotrophic strategies. Protist, 167, 106-120.
Stoecker DK. (1998). Conceptual models of mixotrophy in planktonic protists and some ecological and evolutionary implications. European Journal. Protistology, 34, 281-290.
Stoecker DK, Hansen PJ, Caron DA, & Mitra A. (2017). Mixotrophy in the marine plankton. Annual Review of Marine Science, 9, 311-335.
Villanova V, & Spetea C. (2021) Mixotrophy in diatoms: Molecular mechanism and industrial potential. Physiologia Plantarum, 1-9.