

Emmanuel Romero1, Amelia Sánchez-Pérez2 y Laura Sánchez-Velasco2
1Universidad Autónoma de Baja California Sur, Departamento Académico de Sistemas Computacionales, Carretera al sur km 55, Col. Mezquitito, La Paz, BCS, Mexico
2Instituto Politécnico Nacional-Centro Interdisciplinario de Ciencias Marinas, Departamento de Oceanología, Av. Instituto Politécnico Nacional s/n, Col. Playa Palo de Santa Rita, C.P. 23096, La Paz, Baja California Sur, Mexico
El océano se encuentra en constante movimiento, las corrientes marinas forman enormes remolinos que influyen en la vida marina de maneras sorprendentes. Los remolinos, en específico los de “mesoescala”, pueden llegar a medir de 100 a 300 km, escalas de profundidad de ∼500–1000 m (Klein et al., 2019) y perdurar durante meses (Chelton et al., 2011).
En el hemisferio norte, los remolinos pueden clasificarse como remolinos ciclónicos (Figura 1a), si giran en contra de las manecillas del reloj, estos levantan agua fría y rica en nutrientes del fondo hacia la superficie. Mientras que, los remolinos anticiclónicos (Figura 1b), giran a favor de las manecillas del reloj y hunden agua cálida de la superficie hacia el fondo (Bakun, 2006). Ambos tipos de remolinos influyen en la regulación de la absorción y redistribución de calor (He et al., 2024), el transporte de nutrientes (Sadighrad et al., 2021), la compresión de hábitats (Lachkar et al., 2016) y la distribución de zooplancton (Sánchez-Velasco et al., 2013), entre otros factores. Las larvas de peces, en su fase temprana, forman parte del zooplancton y pasan por tres fases principales (estadios larvarios): (i) preflexión, cuando se desarrollan las partes esenciales del cuerpo; (ii) flexión, cuando se desarrollan las aletas; y (iii) postflexión, cuando se completan los procesos iniciales de crecimiento y las larvas se están preparando para la fase juvenil (Peña et al., 2023).
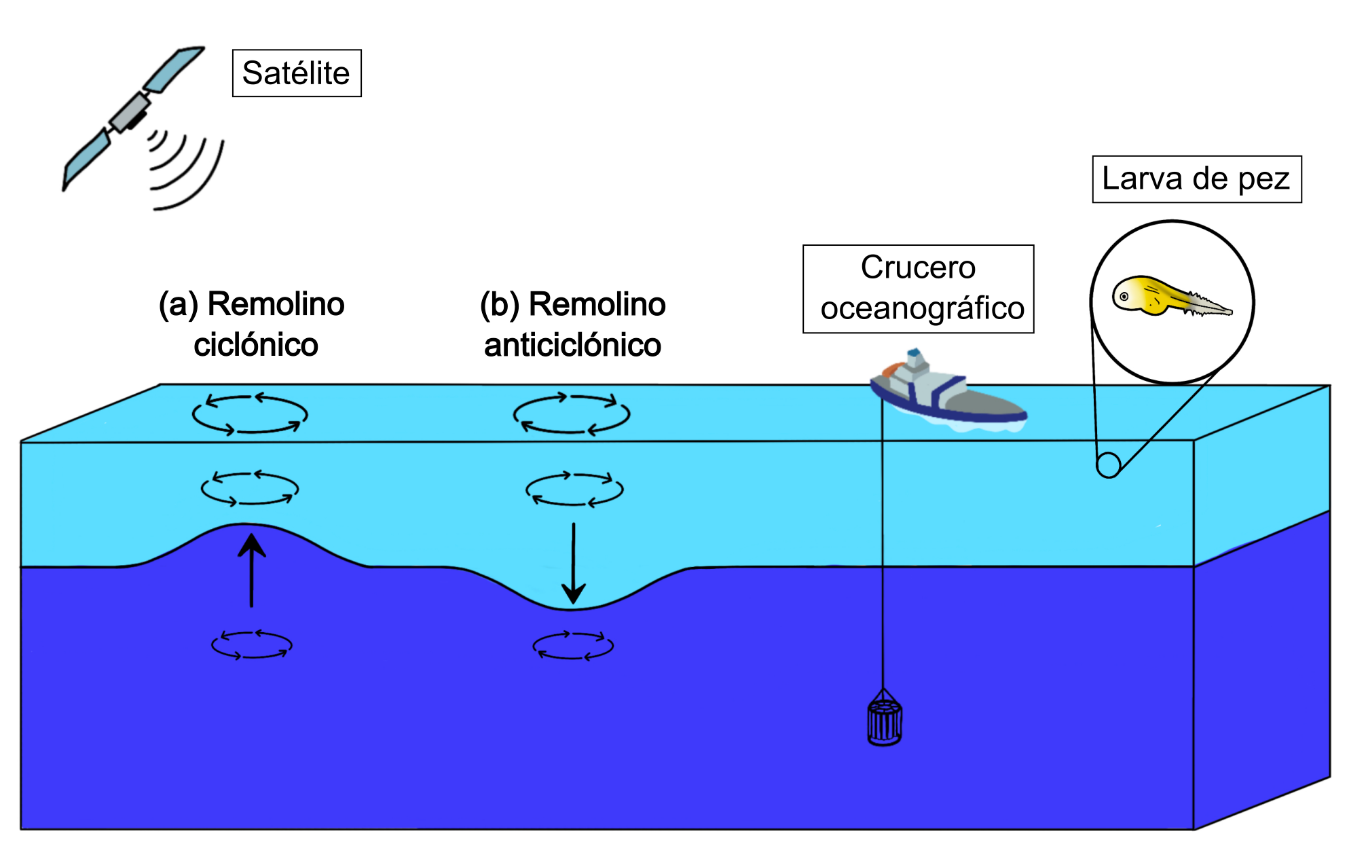
Figura 1. Diagrama esquemático del impacto de los remolinos (a) ciclónico y (b) anticiclónico en el océano.
Un conjunto de estudios recientes ha analizado cómo estas estructuras afectan la distribución de larvas de peces en la entrada al Golfo de California. Durante una expedición oceanográfica en octubre de 2022, Romero et al. (2024) a través de imágenes satelitales identificaron tres grandes remolinos en la entrada al Golfo de California (Figura 2). El crucero oceanográfico sólo pudo atravesar a dos de ellos (A2 y C1) y medirlos en detalle con el uso de un equipo llamado CTD, el cual mide la temperatura, la presión y la conductividad (salinidad). Los resultados revelaron diferencias importantes en la forma en que cada tipo de remolino influyó en la distribución de las larvas de peces. Estas larvas fueron capturadas utilizando una Multinet con sensor de profundidad, la cual descendió con tres redes cerradas y se realizaron arrastres, abriendo las redes a distintas profundidades para para efectuar la captura en áreas específicas dentro de los remolinos.
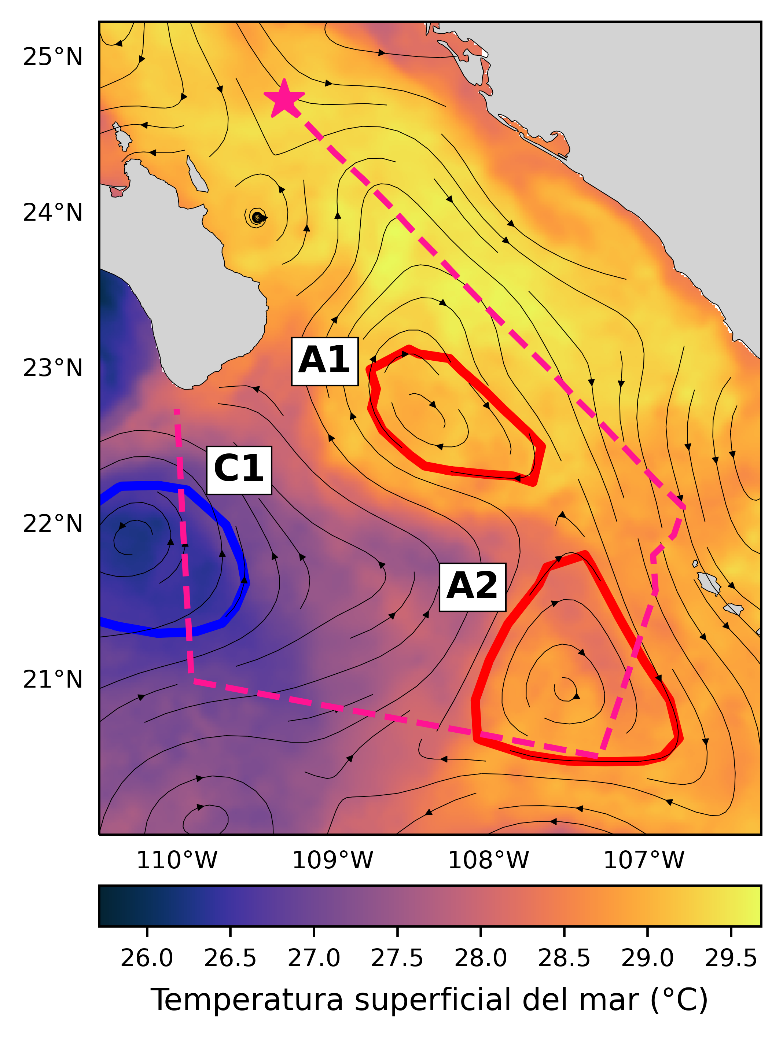
Figura 2. Temperatura superficial del mar (barra de color) con el flujo de las corrientes (flechas negras) del 5 al 20 de octubre de 2022. Se indican dos remolinos anticiclónicos (A1 y A2) en rojo y un remolino ciclónico (C1) en azul. La línea punteada muestra la ruta del crucero oceanográfico, el inicio se marca con una estrella.
El remolino anticiclónico, tenía un mes de haberse formado y medía 15,200 km², generó un hundimiento de aguas cálidas de la superficie hasta alrededor de los 100 metros de profundidad. Este proceso elevó la temperatura y la cantidad de oxígeno disuelto debajo de la superficie del mar. Este remolino se asoció con la mayor concentración de larvas de peces desde aproximadamente 200 m de profundidad hasta la superficie, observándose el predominio de larvas en fase de flexión. Se observó, además, que el remolino retuvo y transportó larvas de dos especies costeras, al Pez linterna (Benthosema panamense) y al Bacalete del Pacífico oriental (Bregmaceros bathymaster).
Por otro lado, el remolino ciclónico, fue el más grande y el más antiguo, midiendo 26,000 km² y formándose nueve meses atrás. Este remolino presentó condiciones opuestas: aguas más frías y menos oxígeno, debido a que levantó agua desde los 530 m de profundidad. En él, se observó una baja abundancia de especies de larvas de peces. Estos hallazgos resaltan cómo los remolinos pueden afectar la distribución de las larvas de peces, sugiriendo que los remolinos anticiclónicos atrapan y retienen a las larvas de peces, mientras que los ciclónicos, las desplazan fuera de su centro.
Con el objetivo de entender de mejor manera cómo los remolinos afectan la distribución de las larvas de peces en la entrada del Golfo de California, Sánchez-Velasco et al. (2025) analizaron datos de tres expediciones oceanográficas, realizadas durante los años 2012, 2015 y 2022. Se Ellos utilizaron modelos computacionales para simular el transporte de estas criaturas diminutas y sus resultados mostraron que, los remolinos de mesoescala actúan como trampas que retienen las larvas dentro de sus estructuras. Sin embargo, el comportamiento varió según el período estudiado.
En abril de 2012, el 85% de las larvas se mantuvieron dentro del remolino durante la primera semana, y solo el 34% logró moverse hacia el norte en la segunda semana. En junio de 2015, más del 90% de las larvas fueron transportadas hacia el norte debido a la Corriente Costera Mexicana. Por último, en octubre de 2020, ninguna larva logró avanzar hacia el interior del Golfo de California. Se concluyó que la alta actividad de los remolinos en la entrada del Golfo de California actúa como una barrera, limitando la dispersión de las larvas y otros organismos planctónicos. Esto sugiere que fenómenos similares podrían estar ocurriendo en otros mares semicerrados del mundo, influyendo en la biodiversidad marina y en las pesquerías.
Palabras clave:
Remolinos, larvas de peces, Golfo de California
Referencias:
Bakun, A. (2006). Fronts and eddies as key structures in the habitat of marine fish larvae: Opportunity, adaptive response and competitive advantage. Scientia Marina, 70(S2), Article S2. https://doi.org/10.3989/scimar.2006.70s2105
Chelton, D. B., Schlax, M. G., & Samelson, R. M. (2011). Global observations of nonlinear mesoscale eddies. Progress in Oceanography, 91(2), 167-216. https://doi.org/10.1016/j.pocean.2011.01.002
He, Q., Mo, D., Zhan, W., Cai, S., Tang, S., Zha, G., & Zhan, H. (2024). Thermal Imprints of Mesoscale Eddies in the Global Ocean. Journal of Physical Oceanography, 54(9), 1991-2009. https://doi.org/10.1175/JPO-D-23-0226.1
Klein, P., Lapeyre, G., Siegelman, L., Qiu, B., Fu, L.-L., Torres, H., Su, Z., Menemenlis, D., & Le Gentil, S. (2019). Ocean-Scale Interactions From Space. Earth and Space Science, 6(5), 795-817. https://doi.org/10.1029/2018EA000492
Lachkar, Z., Smith, S., Lévy, M., & Pauluis, O. (2016). Eddies reduce denitrification and compress habitats in the Arabian Sea. Geophysical Research Letters, 43(17), 9148-9156. https://doi.org/10.1002/2016GL069876
Peña, R., McGregor-Bravo, C. U., & Contreras-Olguín, M. (2023). Allometric growth and larval development in Pacific red snapper Lutjanus peru under culture conditions. Journal of Fish Biology, 102(2), 413-425. https://doi.org/10.1111/jfb.15279
Romero, E., Sánchez-Velasco, L., Tenorio-Fernandez, L., Jiménez-Rosenberg, S. P. A., Sánchez-Pérez, A., Fundora-Pozo, M., & Beier, E. (2024). Mesoscale activity in the northeastern pacific water mass convergence region and its influence on fish larvae distribution by development stages (October 2022). Deep Sea Research Part I: Oceanographic Research Papers, 204, 104228. https://doi.org/10.1016/j.dsr.2023.104228
Sadighrad, E., Fach, B. A., Arkin, S. S., Salihoğlu, B., & Hüsrevoğlu, Y. S. (2021). Mesoscale eddies in the Black Sea: Characteristics and kinematic properties in a high-resolution ocean model. Journal of Marine Systems, 223, 103613. https://doi.org/10.1016/j.jmarsys.2021.103613
Sánchez-Velasco, L., Lavín, M. F., Jiménez-Rosenberg, S. P. A., Godínez, V. M., Santamaría-del-Angel, E., & Hernández-Becerril, D. U. (2013). Three-dimensional distribution of fish larvae in a cyclonic eddy in the Gulf of California during the summer. Deep Sea Research Part I: Oceanographic Research Papers, 75, 39-51. https://doi.org/10.1016/j.dsr.2013.01.009
Sánchez-Velasco, L., Montes-Arechiga, J., Romero, E., Ruvalcaba-Aroche, E. D., Godínez, V. M., Tenorio-Fernández, L., Beier, E., & Ladah, L. B. (2025). Modeling mesoscale circulation and potential fish larvae transport at the entrance of a semi-enclosed sea under different periods. Deep Sea Research Part I: Oceanographic Research Papers, 216, 104430. https://doi.org/10.1016/j.dsr.2024.104430